
我国已成为世界饲料生产大国,但我国饲料资源存在严重缺乏,饲料原料对进口的依存度非常大。2012年我国进口大豆5838万吨,同比增长 11.2%。2012年全国玉米产量为20812万吨,增产1534万吨,全年进口玉米520.74万吨,同比增长197.08%,其中累计进口美国玉米511.33万吨,同比增长20.3%。
随着饲料工业的发展,饲料用粮对玉米、大豆等粮食的刚性需求量将会逐年增加。针对饲料资源紧缺现状,我国“十二五”发展目标提出“坚持开源节流,优化饲料资源配置”。始终把资源开发和高效利用作为保障饲料工业持续发展的根本要求;因此,为缓解玉米紧张的局面,急需开发非常规饲料资源如,大麦、小麦、燕麦、黑麦等代替玉米,以缓解玉米市场的供需矛盾。
本文主要对大麦的品种特性、饲料营养价值、抗营养因子组成及大麦作为饲料开发利用的方法进行概述,以为很好的开发利用大麦能量饲料资源提供指导。
1、大麦的品种特性与营养价值差异
1.1品种特性
大麦是一、二年生草本植物,在植物学分类上属禾本科-大麦属。植株似小麦;比较软叶片略厚而短,颜色淡,叶舌、叶耳较大,无毛;穗状花序,穗轴各节着生三个小穗,每小穗小花一朵;护颖细长呈针状:无芒或具长芒、钩芒;子实扁平,中宽两边较尖,于紧密结合。大麦一般可分为二棱、四棱、六棱大麦。两种大麦杂交后产生一种四棱大麦可以提供相同的营养,但质量要差些。有经济价值的是普通大麦中的两个亚种,即二棱大麦亚种和多棱大麦亚种,通常我们将多棱大麦叫六棱大麦。二棱大麦穗轴每节片上的三联小穗,仅中间小穗结实,侧小穗发育不全或退化,不能结实。二棱大麦穗粒数少,籽粒大而均匀。六棱大麦穗轴每节片上的三联小穗全部结实。一般中间小穗发育早于侧小穗,因此,中间小穗的籽粒较侧小穗的籽粒稍大。两棱大麦具有丰满的内核和更高的容重,淀粉含量要高于六棱品种,平均的营养成分组成仅有一些轻微的差异。两棱大麦通常更适合干燥生长环境,含有较多的蛋白质和酶。高蛋白质的大麦适合作为食品或饲料,含较多酶的大麦麦芽可用来酿酒。在饲喂动物方面,两棱和六棱大麦品种上的差异并不能提供明显的在饲喂动物上的差异。
1.2能量和蛋白
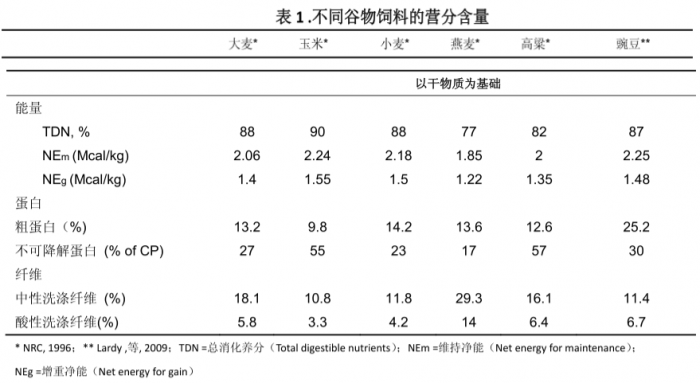
大麦籽粒中含粗蛋白为 11.7%~14.2%,这一数字在谷类籽实中是比较高的,略高于玉米,而与小麦和燕麦相似,但低于豌豆。氨基酸中除亮氨酸 (0.87%) 和蛋氨酸0.14%)外,均较玉米含量高,赖氨酸含量 (0.44%) 接近玉米的 2 倍,但利用率低于玉米。大麦的能量值稍低于玉米和小麦,原因是由于其纤维含量较高 (中性洗涤纤维,酸性洗涤纤维)。大麦是育肥肉牛和肥育猪获得白色朋体所需的良好能量饲料,是肉牛日粮中主要的能量和蛋白质来源,其养分含量 (表1) 并不逊色与玉米、燕麦、小麦、高梁和豌豆。
1.3矿物质和维素含量
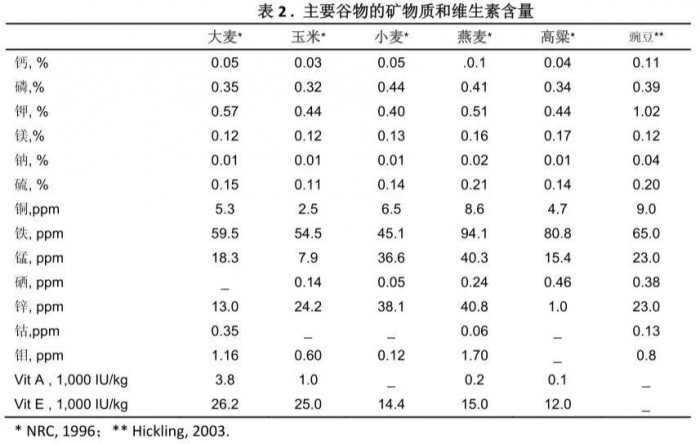
所有谷物的钙含量相对要低,而磷含量要高些(表2)。因此,在高谷物型日粮中有必要补充钙。大麦中的磷含量类似与玉米和高梁,但低于小麦或燕麦。大麦中钾的含量要比其他谷物饲料高。维生素A和E也要高于其它谷物。
1.4淀粉
谷类的主要成分为淀粉。大麦淀粉含量通常在52%-60%之间,但田间生长条件会影响到籽粒淀粉含量和淀粉的自然属性(Myllarinen等,1998 )。在同一地点种植的品种,随着天气的变化(Holtekjglen等,2008)每年会有不同的淀粉成分。随着大麦育种的发展,二棱大麦比六棱大麦含有更多的淀粉和少的纤维。因此,二棱大麦一般比六棱大麦的能量要高出1%-5%大麦含有直链淀粉和支链淀粉的两种主要类别。
“正常”的大麦品种含有约27%的直链淀粉和73%的支链淀粉,而“糯大麦”品种具有较低的直链淀粉(2%-10%)和较高的支链淀粉(90%-98%)的含量(Anker-Nilssen 等,2006)。一些糯大麦品种已被证明含有100%的支链淀粉 (Copeland 等,2008)。
1.5脂肪和其它成分
与玉米、燕麦相比,大麦中脂类含量相当低,仅为2%~3%(Fedak和 Dakoche,1997),其中绝大部分是甘油三醋,其脂肪酸部分主要为软脂酸和不饱和脂肪酸。大麦灰分含量为2.0%~3.0%主要是钾和磷,另有少量的氯、镁、硫、钠、钙、铁、锌和锡。大麦也含有丰富的B族维生素,而基本上不含有胡萝卜素和维生素A、D、K。
2大麦中的抗营养因子
2.1呕吐毒素
呕吐毒素,脱氧雪腐镰刀菌烯醇 (Deoxynivalenol,DON),是一种常见的由禾谷镰刀菌分泌的物质。作为单端抱霉烯族的一员,呕吐毒素一般由寄生在小麦、玉米、大麦与秣草等谷类产品上的真菌生物所产生。呕吐毒素的毒性影响包括:恶心(呕吐)、拒绝进食、肠胃炎、痢疾、免疫抑制与血液病。DON可导致猪采食量减少,呕吐,但并没有证据表明DON对奶牛、育肥牛会产生不良的影响。
初产青年母牛饲喂内含浓度36.8PPM的DON打捆大麦,怀孕期间和哺乳期间分别饲喂8磅/头/天和12磅/头/天,并没有观察到对牛的负面影响。肥育牛饲喂被DON感染的大麦,浓度高达12.6ppm并不影响牛的生产性能或胴体性状。生长和育肥牛饲喂被DON感染的大麦,浓度高达21ppm时,并不会对牛的生长性能和胴体品质产生负面影响(Dicostanzo等1995)。
2.2非淀粉多糖
大麦中非淀粉多糖(Non-Starch Polysaccharides,NSP)主要包括阿拉伯木聚糖和β-葡聚糖,含量分别为3.3%和7.6%,要高于玉米的0.3%和 3.3%。β-葡聚糖包括(1,4)-β-葡聚糖,(1,3)-β-葡聚糖,(1,3-1,4)-β-葡聚糖。
研究表明以大麦为主的日粮粘度增加是由于水溶性葡聚糖所致,也可能是可溶性阿拉伯木聚糖。阿拉伯木聚糖是一种聚戊糖,是阿拉伯糖和木糖的五碳糖分子组成的多糖。大麦胚乳细胞壁中含有20%的戊聚糖。Henry(1987)对2种大麦品种检测结果表明阿拉伯糖含量平均为2.20%,木糖平均为 5.29%。小麦中阿拉伯木聚糖含量相对较低。
与其它谷物相比,大麦中的阿拉伯木聚糖的量低于黑麦,但高于燕麦,高粱,或大米。无芒大麦阿拉伯木聚糖含量显着低于二棱或六棱大麦。六棱大麦品种的阿拉伯木聚糖水平一般比二棱品种略高。β-葡聚糖含量相同的情况下,糯性基因的有无并不对阿拉伯木聚糖的含量产生影响。
经过对大麦无芒基因和糯性基因的遗传选择,以选育出更适合于人类所需的大麦品种(Xue 等,1997)。研究已表明,提高可溶性膳食纤维,特别是 B-葡聚糖,可以改善人类健康。NSP在人类的饮食中已被证明对肠道健康有正面的影响,并可减少冠状动脉心脏疾病,糖尿病和癌症(Holtekjolen等,2008)的发病率。
然而,NSP含量与鸡肠道的粘度、平均日增重、血浆中的总蛋白和低密度脂蛋白胆固醇的浓度、脂类和蛋白质消化率之间是负相关性(Wang等,1992)。雄性肉鸡已被用来作为动物模型研究β-葡聚糖对人类饮食的影响(Wang等,1992)。
2.3单宁
单宁,又称原花青素,或缩合单宁,在动物饲料中使用对生产性能产生负面的效果。大麦中的单宁主要存在于种皮之中。单宁的浓度,因大麦的种类和生长条件而发生变化,一般小于5克/kg干物质。Eggum和 Christensen(1975)分析了29个品种的大麦中单宁含量水平范围为 0.55%至 1.25%。蛋白质消化率的研究表明,大麦单宁含量对蛋白质消化率只有轻微的影响。
2.4植酸
植酸(6-肌醇磷酸)是植物体中磷的主要储藏形式,而磷是动植物生长发育必需的营养元素之一,它不仅是骨骼组织的必需成分,而且对机体其他代谢功能的正常发挥也起重要作用磷是自然界有限的非再生资源。植酸广泛存在于作物种子和果实中。谷物籽实中胚乳、糊粉层富含植酸(Becraft,2001)。植酸与磷结合,减少了家禽对磷的获得量,因此谷物中磷的有效性依赖于磷与植酸结合的水平(Leytem等,2008)。与其它谷物相比,在大麦中植酸的水平低于小麦和燕麦,但高要于黑麦。Salarmoini等(2008) 测量了一些大麦品种中磷的水平。结果表明,尽管测定的样品中总磷水平相似,但植酸含量却表现为显着差异,含量小于0.55%-1.38%。
家禽体内植酸酶的水平是不会有显着变化的,大多数饲料原料中植酸酶的活性也是可以忽略的,但大麦,黑麦,小黑麦,小麦和小麦副产品中的植酸酶有着显着的活性差异。Eeckhout和De Paepe(1994)分析了285个不同样品中植酸酶活性、植酸磷、总磷含量。发现黑麦,小黑麦,小麦和大麦富含植酸酶。大麦中植酸酶的水平和小麦相似,但低于黑麦,高于燕麦在不同大麦品种中,植酸酶水平与总磷,植酸磷的含量并无相关性。因此,不能根据大麦中植酸酶的活性来预测总磷或植酸的含量,谷物中内源性植酸酶的水平是不会影响到家禽生产性能的。
低植酸大麦品种应得到推广应用。研究表明,低植酸大麦品种中磷的生物利用率为49%,而一般大麦仅28%(Li,2001)。低植酸大麦应用于家禽饲料中,可以减少磷的添加量,类便中磷排泄量水平将会降低50%(Salarmoini 等,2008),以减少磷对环境的污染(Jang 等,2003)。
低植酸植物中除了磷的利用率增加外,锌的利用率也会得到提高 (Linares 等,2007)。
3、提高大麦利用率的措施
3.1水侵泡处理
研究已报道将大麦谷物浸泡在水中之后再规划到家禽日粮中饲喂家禽,对家禽的生长性能具有正面的影响。原因可能是由于水溶性的β-葡聚糖得到了去除,或是由于内源性的酶对β-葡聚糖进行了酶解,或两者共同作用的效果。谷物中的化学成分通常是被细胞壁或者其它组分相互隔离的。
例如,碳水化合物和酶解碳水化合物的酶是相互分离的。当相互间的屏障被打开内源性的酶就会作用与碳水化合物。在浸泡的大麦中加入抗生素会抑制水对侵泡大麦营养组分的变化,可能是由于产酶微生物的生长被抑制,不能产生降解酶而使大麦的营养组分发生变化(Thomas 等,1961)。在大多数研究中,浸泡大麦时水的加入量通常按重量计为1份大麦1份水。浸泡时间从30分钟到24小时(Moss等,1982)。大麦经水处理已被证明可以改善肉鸡(Yasar 和 Forbes,1999),生长期蛋公鸡(Lepkovsky和Furuta,1960),幼龄种火鸡(Fry等,1958),日本鹤鸭(Moss 等,1982)的生产性能。Yasar 和Forbes(1999)报道侵泡过的大麦有利于降低肠道粘度,促进增消化道分绒毛层的发育,降低肠上皮隐窝细胞的增殖。
3.2发酵处理
无芒糯大麦与真菌(Rhizopus oligosporus)一起发酵后饲喂肉鸡,可以改善肉鸡的生产性能(Newman等,1985)。大麦被降解β-葡聚糖的乳酸菌发酵后,也可以改善肉鸡生长和早期的饲料转化效率(Skrede 等,2003)。
3.3热处理
热处理后的大麦谷物可提高肉鸡的早期生长性能,但肠道粘度会有所增加 (Gracia 等2003)。大麦经蒸汽处理50分钟,通过滚揉再磨碎。此过程中大麦谷物中淀粉和纤维成分被裂解,促进了酶与营养物质的接触,有利于营养物质被动物的消化吸收。
3.4制粒
Allred 等(1957)报道,大麦制粒可促进新汉夏鸡x白奥海鸡的生长,提高饲料转化效率。Pettersson和Aman(1991) 报道了相似的结果,以大麦为主的日粮经过制粒可以提高日粮的消化率。但也有相反的报道,Ankrah等(1999)研究表明制粒并不会改善生长,提高饲料转化效率,但可降低肠道食糜粘度和提高淀粉的消化率。Arscott等(1958)报道,大麦基础日粮经过制粒,可以促进肉鸡的生长和改善饲料转化效率,但效果要低于以玉米为主的日粮。Sibbald(1976)报道,制粒还可以增加大麦真代谢能。
3.5发芽处理
糊粉层为种子胚乳的最外层组织。在大多数谷类(小麦,黑麦,燕麦,水稻,玉米)中,它是一种单细胞层,但在大麦中是一个多细胞的煳粉层。谷物中胚乳,煳粉层中含有约 30%谷物蛋白质。种子发芽过程中,植物胚胎将会产生赤霉素,这将触发的煳粉层细胞释放出淀粉酶而有助于淀粉的水解,并促进贮藏蛋白发育成胚乳,淀粉胚乳提供糖分而促进根的发育。这种发育过程可以被植物脱落酸抑制,使种子处于休眠状态。
Macgregor和Matsuo(1982)报道,在培养皿中发芽的大麦a-淀粉酶活性将会增加。在发芽48小时内酶合成的速度相对缓慢,48小时后合成速度加快,到164小时后也并未减弱。Peer和Leeson(1985)水培发芽大麦17 天,随着发芽时间的延长,大麦芽代谢能逐渐减少。干燥和研磨豆芽可以提高大麦芽的消化率。发芽在48至96小时,发大麦水取物粘度降低。鸡试验表明,发芽大麦并不影响鸡的生长、饲料转化率,干物质和脂肪的消化将会得到改善。
3.6辐射处理
用伽玛钻60射线照射无芒和有芒大麦后用作鸡饲料,可以提高鸡的生产性能(Campbell等,1986),且辐射剂量和3周龄体重具有曲线相关性。6毫拉德辐射水平时,鸡生产性能的改善达到最高。辐照会使大麦样品中β-葡聚糖的溶解性增加,肠道提取物粘度降低。可溶性的粘性纤维,如大麦中β-葡聚糖,很容易在辐照下发生裂解。同样,AI-Kaisey等(2002)报道了射线照射后的大麦可以减少肠道粘度,辐照处理的大麦种子中β-葡聚糖聚合物的结构可以被打破,从而导致低的β-葡聚糖结构和粘度。
3.7高湿储存
高湿环境下储存(60%干物质)大麦已被证明可以降低β-葡聚糖的水平,提高饲用价值(Svihus等,1997a)。Svihus 等(1997b)报道,在高湿度下,纤维含量水平发生迅速的变化,粘度降低,因为水溶性β -葡聚糖含量的减少,此过程与酶的活性和乳酸菌活性并没有相关性。Aman等(1990)报道,高水分密闭贮存下大麦总的化学成分发生微小变化,但蛋白质、淀粉和 β-葡聚糖中的溶解度却有着显着的变化。然而,这些变化并没有显示出会影响到大麦的营养特性。
高水分贮存饲料大麦是一种可行的替代干燥的方法。Perttila等(2001)研究表明,青增加大麦营养物质的消化率和代谢能。此外,青贮会减少β-葡聚糖的负面影响。在这项研究中所用的青贮大麦收获于成熟阶段,水分含量 42.3%。大麦被与甲酸保存在一起,然后将其存储在小规模的筒仓并压实。大麦在高水分(60%)条件下存储,可以减少抗营养因子的影响,同时高湿度存储导致的真菌污染和相关的真菌毒素的出现也是可能会发生的问题。
3.8添加酶制剂
以大麦为主的日粮中添加酶制剂,可以改善肉鸡的生长性能(Leeson和 Caston,2000:J6zefiak等,2006),来航雏鸡 (Marquardt 等,1994),火鸡(Moran 和 McGinnis,1968)和鸭(Hong 等,2002),但在蛋鸡日粮中补充酶的效果报道并不一致。Brenes等(1993)报道以大麦为主的日粮中添加酶制剂并不影响初始蛋重,平均蛋重,蛋壳质量和死淘率。Francesch等(1995) 报道添加复合酶(β-葡聚糖,木聚糖酶,果胶酶) 不会影响蛋鸡生产或卵子质量,但可增加早期蛋重,减少类便水分含量和脏蛋的数量。
酶制剂对大麦利用影响的效果取决于大麦品种以及大麦种植的环境条件。补充酶制剂是由于大麦胚乳细胞壁中β-葡聚糖的存在。日粮中的β-葡聚糖的水平将会影响酶制剂剂量反应Campbell等(1989)对16个大麦培育品种进行研究,所选择的大麦品种不同之处在于其提取物的粘度不同,这个迹象暗示β-葡聚糖水平将会不同。
雏鸡采食低粘度的大麦要比高粘度大麦品种具有更好的生长性能和饲料转化效率。Campbell等(1993)比较了无大麦和有芒大麦发现了类似的结果。酶制剂的影响效果对有芒大麦比无芒大麦更明显,两种大麦中补充酶要比不补充酶制剂的日粮组生长性能和饲料转化效率具有更好的改善效果。此外,Ravindran等(2007)报道,酶制剂补充对糯大麦要比普通大麦品种具有更好的效果,可能是由于糯大麦品种相对与普通大麦品种含有较多的水溶性β-葡聚糖。
家禽的年龄也是一个需要考虑的因素。家禽在孵化后肠壁结构发生变化,特别是在第7天之后(lji,2001)。Brufau等(1991) 报道,以大麦为基础的颗粒日粮中补充酶制剂,可以改善肉鸡的生长性能和饲料转化效率,然而,在饲养的后期其改善的效果要明显的好于前期。同样,Boguhn 和 Rodehutscord(2010)报道以谷物为基础的火鸡日粮(小麦、大麦、黑麦),添加酶制剂可以改善生长和饲料转化效率,尤其是在后期的生长阶段。Salih 等(1991)报道日龄大的鸡似乎能更好地应对高粘度的大麦。
大麦日粮中酶制剂的有效性也受家禽类型的影响。Rotter等(1989)报道,在大麦基础日粮中添加酶制剂对肉鸡和来行雏鸡产生相似的影响效果。肉鸡对不补充酶制剂的大麦日粮所带来的负面效应要更敏感。Sott等(2001) 报道,来杭鸡喂大麦基础日粮其肠道粘度是肉鸡的两倍。
补充商业性饲料酶可以增加家禽对日粮中植酸磷的利用率。非植酸磷水平低的肉鸡大麦基础日粮中补充植酸酶可增加磷的沉积,减少类便中磷的排泄量可达到45%(Juanpere等,2004)。植酸也可以结合蛋白质,使消化酶不能作用于蛋白质。已报道在家禽日粮中补充植酸酶可以增加氨基酸的可利用率,但增加的程度有限(Bedford等,2000)。Juanpere等(2005)和 Cowieson等(2005) 报道在大麦基础日粮中添加植酸酶和β-葡聚糖具有相加效应目前,已选育出了转基因大麦品种。转基因大麦可以产生耐热性的芽抱杆菌葡聚糖酶(VonWettstein等,2000),是一种沉积在谷物蛋白体内的酶。肉鸡日粮中添加麦芽(发芽)的转基因大麦可以改善生长和饲料转化效率,减轻但不能完全消除粪便粘性的现象。转基因大麦内含大量的高活性β-葡聚糖酶。将转基因大麦添加到饲料日粮中可以起到与酶制剂相同的作用。Horvath等(2001)用转基因麦(4.28g,葡聚糖/g总可蛋白)添加到大麦料中饲喂幼鸡,结果发现幼鸡增重明显。幼鸡小肠和排泄物中可溶性葡聚糖的含量比对照组分别降低了75%和50%。排泄粘性物质的幼鸡数量显着减少 (Wettstein等,2000)。
4、小结
饲料资源紧缺的情况下,为了缓解主要能量饲料玉米的供需矛盾,大麦、小麦、黑麦是良好的玉米替代品。大麦中含有呕吐毒素、非淀粉多糖、丹宁、植酸等抗营养因子。经过水侵泡处理、发酵处理、高湿度贮存、制粒、辐照处理、发芽处理、添加酶制剂等措施可以使大麦得到很好的利用。
家禽饲料中添加大麦并补充β-葡聚糖酶,可作为家禽能量饲料的来源。日龄较大的家禽比幼雏能更好的利用大麦。添加大麦所补充酶的水平将取决于家禽的年龄,以及所使用的大麦品种,生长条件,收获时期和贮存时间。
家禽日粮中添加转基因大麦可以改善生长性能和饲料转化效率,减轻但不能完全消除类便粘性的现象。植酸酶可以补充到家禽大麦型日粮中提高植酸磷的利用率,以降低对无机磷的补充。大麦基础日粮中添加植酸酶,β-葡聚糖酶具有相加效应。
(文章来源:中粮营养健康研究院;作者:王勇生)