油脂效率、蛋白吸收 – 溶血磷脂助您“一网打尽”
油脂效率、蛋白吸收 – 溶血磷脂助您“一网打尽”
建明水产科技倾心之作 – 水产利舒宝营养吸收强化剂
2022年水产行业热点话题已经从如何替代鱼粉减少油脂用量变成了如何有效替代豆粕并减少油脂用量,可想而知当前饲料企业所面对的原料成本上涨压力。2020下半年开始豆油价格勇往直前,目前维持在1万元/吨左右。2022短短几个月时间,豆粕价格最高飙升超过5000元/吨。水产动物对于碳水化合物的利用非常有限,所需营养素和能量主要来自脂肪和蛋白。因此改善油脂利用,强化蛋白吸收可以有效地缓解部分成本压力,同时助力水产企业优化饲料配方。
第一部分
脂类和蛋白质是水生动物的主要有机成分,它们在水生动物生长(包括繁殖)和运动(包括迁徙)方面发挥着重要的代谢能量来源作用,其中脂类占主导地位。从极性上分,脂类可分为两大类:极性脂类和中性脂类。极性脂类以磷脂为主,为膜结构的主要成分(图1);中性脂类以甘油三酯为主,为动物主要的能量来源(图2)。水生动物体内的脂类成分中,饱和脂肪酸和单不饱和脂肪酸优先酯化于sn1和sn3位置,而多不饱和脂肪酸(PUFA)优先酯化于sn2位置(图1)。磷脂也有相类似结构特征,sn3位置由磷酸基团取代,饱和脂肪酸和单不饱和脂肪酸优先酯化于sn1位置,多不饱和脂肪酸PUFA优先酯化于sn2位置(图2)。其他脂类还包含蜡酯(单个脂肪酸和单个脂肪醇酯化)、鞘脂类和胆固醇(Tocher et al., 2003)。溶血磷脂是在磷脂的基础上加工得到的,磷脂的结构包含一个磷酸基团和两个脂肪酸,溶血磷脂的结构则包含一个磷酸基团和一个脂肪酸。原料来源加工工艺不同所得溶血磷脂结构会有很大的差异。

图1 甘油三酯结构:三个脂肪酸分别酯化在sn1、sn2和sn3位置
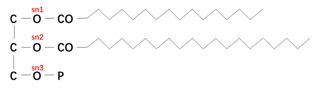
图2 磷脂结构:两个脂肪酸酯化于sn1和sn2位置,磷酸基团位于sn3位置
脂类在水产饲料配方中有举足轻重的地位,一方面提供了生长繁殖所需的能量和必须脂肪酸,同时作为脂溶性维生素和色素的载体增强其吸收利用率,并优化水产饲料的适口性。其主要生理功能如下:
能量供应
a) 能量供应
油脂(甘油三酯)在水生动物体中最重要的作用包括 ①脂肪的存储 ②通过脂肪酸β氧化并以ATP的形式满足动物体代谢所需能量(Sargent et al, 1989; Froyland et al., 2000)。
b) 繁殖
不仅为代谢提供能量,脂质也为水产动物繁殖提供所需的能量(Henderson et al., 1984b)。Marshall et al., 1999阐述了鳕鱼繁殖能力和油脂极强的关联度。
c) 胚胎发育和幼体发育
不同品种胚胎和幼体发育所需的脂肪酸组成差异较大。整体上,海水鱼类鱼卵极性脂质含量较高(60-90%),淡水鱼类鱼卵中性脂质含量较高。但也有少数例外,例如海鲈鱼卵中中性脂质比较超过50%(Ronnestad et al., 1998)
膜结构和膜功能
磷脂双分子层是构成细胞膜的基本支架,居间镶嵌其他成分包括糖蛋白、糖脂、蛋白质和胆固醇等。其中磷脂占总量约70%。
其他功能
a) 信号传递:固醇类激素的合成
b) 糖蛋白、糖脂:参与信号识别和免疫
c) 其他激素和色素的前体
水产养殖过程中关注最多的是脂肪的存储和供能,简单来说对于鱼苗到成鱼的养殖过程中,高脂饲料对于动物肝脏的压力越小越好,肝脏中脂肪分解为脂肪酸的β-氧化供能则多一些更好。缓解肝脏压力,强化脂肪酸氧化供能,参见下文溶血磷脂对水产动物肝脏脂肪代谢酶活的影响(图3,图4)。
水产动物脂肪代谢过程(图3)
水产动物采食之后,配方中不论是中性的甘油三酯还是极性的磷脂,不可或缺的第一步都是脂类的乳化。胆汁酸(盐)的分泌可以帮助适量油脂在肠道中的乳化。随着高油脂高能值水产配方的普适性,有必要额外添加一些元素以促进油脂的乳化,例如植物来源的溶血磷脂。相同实验条件下(表1),可以看到溶血磷脂显著提高了同一底物不同脂肪酸的释放率(p<0.05)。油脂乳化中另外一个需要关注的是形成乳糜微粒所需乳化剂的数量,也就是临界胶团浓度(CMC)。其定义是指形成乳糜微粒所需的乳化剂或表面活性剂的最低浓度,浓度越低性能越好。表2中可以看出,只需要0.02-2微摩尔的溶血磷脂就能形成乳糜微粒。
表1 相同条件下,不同元素对脂肪酸释放量的影响(Kemin internal data)
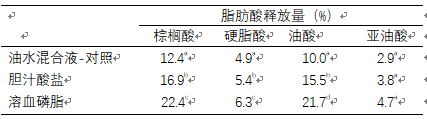
表2 不同成分临界胶团浓度(Biochemistry, 1984)

乳化之后就到了脂肪酶的出场时间了,脂肪酶主要是由胰腺(肝胰腺)分泌,也有研究表明肠道中小部分脂肪酶可能由肠粘膜分泌而来(Fange et al., 1979; Smith, 1989a)。不过最终目的都是一致的,将甘油三酯分解为甘油单酯和游离脂肪酸(图3,第2步),并以自由扩散的方式进入肠道上皮细胞。进入到肠道上皮细胞内的甘油单酯、游离脂肪酸、少部分甘油双酯和甘油会在内质网上完成再酯化反应,重新合成甘油三酯(Tocher et al., 2003)。肠道上皮细胞内,再酯化的甘油三酯和胆固醇以及载脂蛋白依据脂肪酸的不同类型分别形成乳糜微粒或极低密度脂蛋白(VLDL)(图3,第3步)。所形成的乳糜微粒或VLDL主要通过淋巴系统经内循环进入肝脏。
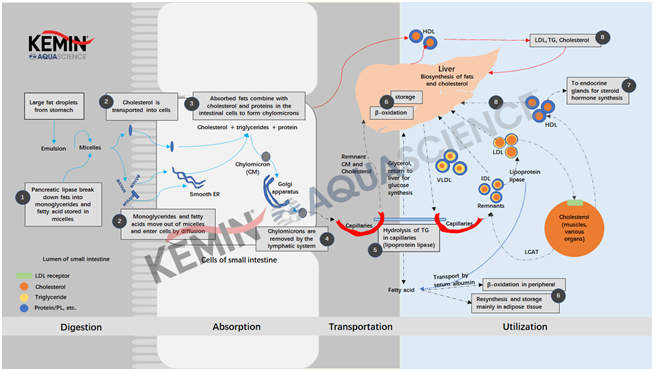
图3 水产动物油脂(甘油三酯)代谢过程,包括脂肪的消化、吸收、运输和利用。
乳糜微粒在进入到肝脏之前有极少部分会被脂蛋白脂酶分解供机体使用。大部分的油脂进入到肝脏经过分解和再酯化的过程,分解的部分脂肪酸通过肉碱棕榈酰转移酶1(CPT1)转移到线粒体中β氧化提供能量(图3第6步)(Tocher et al., 2003);肝脏中再酯化合成的甘油三酯部分存储在肝脏中,部分通过VLDL的形式运输到外周组织。为了达到更好的生产性能,大多数水产动物的配方能值偏高,因此提高肝脏对于脂肪的处理能力至关重要。恰当合理的脂肪沉积有助于缓解脂肪肝的压力,同时提高CPT1酶活力可以有效强化脂肪酸的β氧化供能。合理配比的溶血磷脂复合产品(水产利舒宝)可以有效的促进CPT1酶活(图4),并优化脂肪在肝脏中的存储(相同实验中,肝脏油红切片数据清晰展示该结果)。
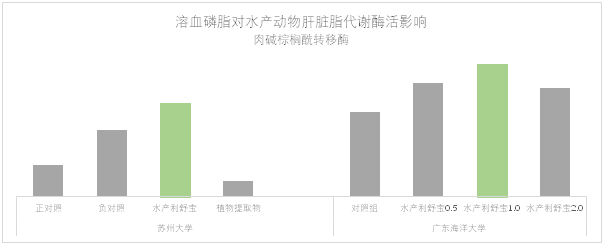
图4 溶血磷脂对水产动物肝脏脂代谢酶活影响(TD-21-7292)
转运出肝脏的VLDL,在外周组织中被脂蛋白脂酶和肝脂肪酶逐步分解,酶解出的脂肪酸 ①用于外周组织氧化供能 ②再合成并存储于脂肪组织中(图3,第6步)。酶解剩余部分可直接被肝脏重新吸收,或者进一步被脂蛋白脂酶和肝脂肪酶分解而得到中间密度脂蛋白IDL和低密度脂蛋白LDL。另外一部分VLDL酶解剩余产物可直接形成“初级高密度脂蛋白”,肝脏自身也能合成“初级高密度脂蛋白”。血液中的“初级高密度脂蛋白”从外周组织中捕捉游离胆固醇,并通过磷脂胆固醇酰基转移酶(lecithin-cholesterolacyltransferase,LCAT)催化脂蛋白中磷脂的sn2位脂肪酸转移给游离胆固醇,生成胆固醇酯,并形成“成熟高密度脂蛋白”(Babin et al., 1989)。高密度脂蛋白(图3,第8步)不仅可以清除多余的游离胆固醇,因其颗粒度小(7.5-12.5纳米)可自由进出血管管壁,并摄取血管壁内膜底层沉积下来的低密度脂蛋白和甘油三酯等“有害物质”,转运到肝脏进行分解代谢(张会琴等,2010)。
高密度脂蛋白就像一辆运载能力很强的迷你卡车,可以将血管中不该存在的胆固醇、甘油三酯和低密度脂蛋白运回肝脏。一方面改善了水产动物的健康程度,另一方面将脂质运回肝脏提高“废物利用率”。研究表明配方中添加溶血磷脂(水产利舒宝)可以有效提高机体高密度脂蛋白含量(图5)。
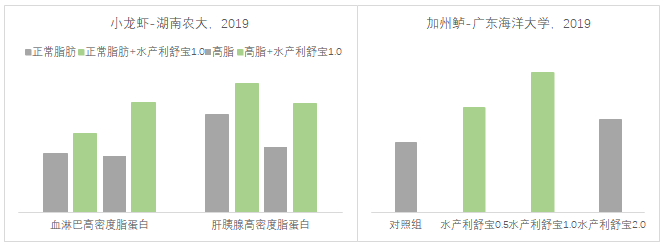
图5 不同配方脂肪溶血磷脂对小龙虾机体高密度脂蛋白(mmol/L)的影响;溶血磷脂对加州鲈血清高密度脂蛋白的影响
第二部分
油脂的消化吸收离不开乳化作用以增大乳液胶团微粒的比表面积,从而扩大脂肪与脂蛋白脂酶和肝脂肪酶的接触,并加速酶解反应。蛋白质的消化吸收离不开蛋白本身空间结构的变化(例如在胃酸的作用下蛋白质三维结构打开成长肽链),同时降低蛋白微滴的颗粒度可以增大蛋白质与蛋白酶(胃蛋白酶、胰蛋白酶、外源添加的符合水产动物肠道的碱性蛋白酶)的接触面积。
恰当的外源功能性营养物质的添加可以有效降低蛋白微滴的颗粒度,例如磷脂和溶血磷脂(Mine et al., 1992, 1993)。Ohtsuru等在1976年发现,大豆磷脂和大豆蛋白通过磷脂疏水端与蛋白疏水端直接结合和蛋白表面平铺磷脂层两种方式连接在一起。随后Dutilh等在1981年发现蛋黄脂蛋白(脂蛋白主要成分之一为磷脂)在与磷脂酶反应之后,其乳化性能和稳定性有了极大的提升。
基于以上研究,Mine等于1992、1993年,针对磷脂和溶血磷脂对蛋白(鸡蛋中提取的卵清蛋白)微滴的粒径做了评估和研究,结果发现卵清蛋白平均微滴粒径为6.6微米,磷脂则改善了粒径到2.4微米,溶血磷脂极大的改善了微滴粒径到0.7微米。研究中发现溶血磷脂对于蛋白微滴粒径的改变是因为对蛋白α-螺旋和β-结构的改变(Mine et al., 1992, 1993)。因此可以看到溶血磷脂可以改善蛋白微滴粒径,增大其比表面积,扩大蛋白和蛋白酶的接触面。
蛋白主要以氨基酸和小肽的形式被肠道上皮细胞吸收,和甘油单酯/脂肪酸的自由扩散方式不同,氨基酸和小肽的吸收需要相应的转运载体。溶血磷脂本质上为多种溶血磷脂酰复合物,其中多个活性成分可有效激活并提高肠道蛋白质相关转运载体的基因表达,如小肽转运载体Pept1及氨基酸转运载体CAT2等。动物实验同样验证了该结论,配方中蛋白含量减少2%之后添加0.1%的水产利舒宝可以充分提高蛋白的吸收,实验组蛋白存留率为30.38%,高出对照组3.73%,同时不影响生长性能(图6)。
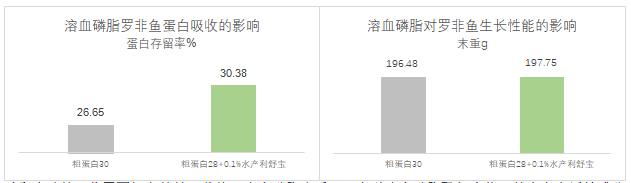
图6,溶血磷脂对罗非鱼蛋白吸收和生长性能的影响
第三部分
优化成本有多种方案,建明水产科技为您保驾护航:
① 配方调整,提高油脂消化吸收和利用率(建明水产科技:水产利舒宝)
溶血磷脂有效提高了脂肪酸释放量(表1),且形成乳糜微粒所需的量最少(表2),
溶血磷脂缓解了高脂配方带来的肝脏压力,强化了脂肪酸β-氧化供能(图4),
溶血磷脂优化了脂肪内循环和“废物”利用率并改善了动物的健康程度(图5);
② 原料优化,强化蛋白效率(建明水产科技:水产利舒宝)
改善蛋白微滴粒径增加比表面积,
提升小肽/氨基酸转运载体基因表达优化蛋白吸收和运输系统
添加符合水产动物肠道需求的外源蛋白酶(建明水产科技:渔酶宝TM液体蛋白酶)。
③ 节能减排,在生产过程中添加工艺助剂(建明水产科技:膨力康TM),以提高生产效率、节省电耗。
详细实验数据及应用方案请咨询建明水产科技的技术专家。
油脂效率、蛋白吸收 – 建明水产科技水产利舒宝营养吸收强化剂助您“一网打尽”。

建明水产科技致力于支持大豆产品的可持续发展
水产利舒宝营养吸收强化剂通过RTRS认证
Reference
Douglas R. Tocher (2003): metabolism and Functions of Lipids and Fatty Acids in Teleost Fish, Reviews in Fisheries Science, 11:2, 107-184
Sargent, J. R. (1989) Ether-linked glycerides in marine animals. In: Marine Biogenic Lipids, Fats and Oils, (Ackman, R. G., Ed.). pp. 175–198.
Froyland, L., O. Lie, and R. K. Berge. Mitochondrial and peroxisomal β-oxidation capacities in various tissues from Atlantic salmon Salmo salar. Aquacult. Nutr., 6: 85–89 (2000).
Henderson, R. J., J. R. Sargent, and B. J. S. Pirie. Fatty acid catabolism in the capelin Mallottus villosus (Muller) during sexual maturation. Mar. Biol. Letters, 5: 115–126 (1984b).
Marshall, C. T., N. A. Yaragina, and Y. Lambert. Total lipid energy as a proxy for total egg production by fish stocks. Nature, 402: 288–290 (1999).
Ronnestad, I., W. M. Koven, A. Tandler, M. Harel, and H. J. Fyhn. Utilization of yolk fuels in developing eggs and larvae of European sea bass (Dicentrarchus labrax). Aquaculture, 162: 157–178 (1998).
Biochemistry, 1984. Ed. G. Zubay. Addison-Wesley
Fänge, R. and D. Grove. Digestion. pp. 161–260. In: Fish Physiology, Vol. VIII. (Hoar, W.S., D. J. Randall, and J. R. Brett, Eds.). New York: Academic Press (1979).
Smith, L. S. Digestive functions in teleost fishes. pp. 331–421. In: Fish Nutrition, 2nd ed. (Halver, J. E., Ed.). San Diego: Academic Press (1989a).
Babin, P. J. and J.-M. Vernier. Plasma lipoproteins in fish. J. Lipid Res., 30: 467–489 (1989).
张会琴,张怀明.《糖尿病预警》:军事医学科学出版社 ,2010:82-83
Mine, Y., Kobayashi, H., Chiba, K. and Tada, M. 31P NMR Study on the Interfacial Adsorptivity of Ovalbumin Promoted by Lysophosphatidylcholine and Free Fatty Acids. J. Agric. Food Chem. 1992, 40, 1111-1115
Mine, Y., Chiba, K. and Tada, M. Effect of Phospholipids on Conformational Change and Heat Stability of Ovalbumin. Circular Dichroism and Nuclear Magnetic Resonance Studies. J. Agric. Food Chem. 1993, 41, 157-161
Ohtsuru, M.; Kito, M.; Takeuchi, Y.; Ohnishi, S. Association of Phosphatidylcholine with Soybean Protein. Agric. Biol. Chem. 1976, 40, 2261-2266
Dutilh, C. E.; Groger, W. Improvement of Product Attributes of Mayonnaise by Enzymic Hydrolysis of Egg Yolk with Phospholipase Az. J. Sei. Food Agric. 1981, 32, 451-458.
作者:建明水产科技技术经理 刘亮 曹杰,未经授权禁止转载